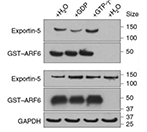
WB蛋白免疫印迹
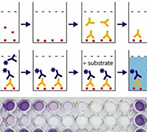
ELISA检测
流式检测
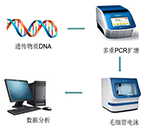
荧光定量PCR(qPCR)实验
核酸提取实验
细胞STR鉴定
多克隆抗体制备
单克隆抗体制备
纳米抗体制备
原核表达系统
真核表达系统
昆虫表达系统
细胞增殖检测
细胞凋亡检测
细胞周期检测
动植物生化分析
小动物核磁检测
小动物CT检测
增强型化学发光检测试剂盒(皮克级)
灵敏化学发光检测试剂盒(飞克级)
通用型抗体稀释液
伊红染液(醇基)
苏木精染液(低浓度)
粪便基因组提取试剂盒
土壤基因组提取试剂盒
唾液DNA提取试剂盒
Human Junctional Adhesion Molecule 3 (JAM3)ELISA Kit
Human Nesprin 2 (Nesp2)ELISA Kit
Human Complement Factor H Related Protein 4 (CFHR4)ELISA Kit
RPMI-1640无糖 (含L-丙氨酰-L-谷氨酰胺,不含酚红)
RPMI-1640无糖 (含L-丙氨酰-L-谷氨酰胺、HEPES)
RPMI-1640无糖(含L-丙氨酰-L-谷氨酰胺)
高效RIPA裂解液(组织/细胞)
0.5%EDTA溶液
0.02% EDTA溶液(Versene液)